Постулаты полинга и кори

В химии, раскраска по CPK (англ. Corey-Pauling-Koltun, рус. Кори-Полинг-Колтун) — популярная схема цветового обозначения атомов различных химических элементов в молекулярных моделях. Данная схема названа в честь молекулярной модели CPK, созданной Робертом Кори и Лайнусом Полингом и усовершенствованной Уолтером Колтуном.
История[править | править код]
В 1952 году, Кори и Полинг опубликовали описание объёмной модели белков и других биомолекул, которую они создали в Калифорнийском технологическом институте.[1]
Их модель представляла атомы деревянными шариками, раскрашенными в яркие цвета, соответствующие химическим элементам. Их цветовая модель включала:
- белый для водорода
- чёрный для углерода
- голубой для азота
- красный для кислорода
Они также построили уменьшенные модели, используя пластиковые шарики такой же цветовой схемы.
В 1965 году Колтун запатентовал улучшенную версию модели Кори-Полинга.[2] В своём патенте он упоминает следующие цвета:
- белый для водорода
- чёрный для углерода
- голубой для азота
- красный для кислорода
- тёмно-жёлтый для серы
- пурпурный для фосфора
- оттенки от светло- до тёмно-зелёного соответственно для галогенов (F, Cl, Br, I)
- серебряный для металлов (Co, Fe, Ni, Cu)
Типичные цветовые обозначения[править | править код]
Типичные цветовые обозначения в CPK включают:
| водород (H) | белый | |
| углерод (C) | чёрный | |
| азот (N) | синий | |
| кислород (O) | красный | |
| фтор (F), хлор (Cl) | зелёный | |
| бром (Br) | коричневый | |
| иод (I) | тёмно-фиолетовый | |
| благородные газы (He, Ne, Ar, Xe, Kr) | голубой | |
| фосфор (P) | светло-оранжевый | |
| сера (S) | жёлтый | |
| бор (B), большинство переходных металлов | бежевый | |
| щелочные металлы (Li, Na, K, Rb, Cs) | фиолетовый | |
| щелочноземельные металлы (Be, Mg, Ca, Sr, Ba, Ra) | тёмно-зелёный | |
| титан (Ti) | серый | |
| железо (Fe) | темно-оранжевый | |
| другие элементы | розовый |
Некоторые из цветов CPK мнемонически связаны с цветами чистых элементов или их соединений. Например, водород — бесцветный газ, углерод в виде угля или графита чёрный, многие формы серы имеют жёлтый цвет, хлор — зеленоватый газ, бром — тёмно-красная жидкость, иод в эфире фиолетовый, одна из аллотропных модификаций фосфора имеет красный цвет, железная ржавчина имеет тёмно-оранжевый оттенок, и т. д. В случае других элементов, таких как кислород и азот, выбор цвета менее понятен. Вероятно, красный цвет для кислорода был выбран потому что он часто ассоциируется с горением, а голубой цвет для азота обусловлен тем, что это главный компонент земной атмосферы, имеющей голубой оттенок.
Современные варианты[править | править код]
Следующая таблица представляет собой цвета, назначаемые каждому элементу в популярных компьютерных программах. Колонка C содержит изначальные цвета Кори и Полинга,[1] колонка K содержит цвета из патента Колтуна.[2] Колонка J описывает цветовую схему, используемую молекулярным визуализатором Jmol.[3] Колонка R описывает цветовую схему, используемую программой RasMol; в случае если указаны два цвета, второй актуален для версий 2.7.3 и выше.[3][4] Все цвета даны приблизительно и могут зависеть от дисплея и настроек видео.
Примечания[править | править код]
- ↑ 1 2 Robert B. Corey and Linus Pauling (1953): Molecular Models of Amino Acids, Peptides, and Proteins. Review of Scientific Instruments, Volume 24, Issue 8, pp. 621—627. doi:10.1063/1.1770803
- ↑ 1 2
Walter L. Koltun (9165), Space filling atomic units and connectors for molecular models. U. S. Patent 3170246. - ↑ 1 2
Jmol color table на sourceforge.net. - ↑
Rasmol color table Архивировано 13 мая 2001 года. на bio.cmu.edu.
Ссылки[править | править код]
- What is the basis for the CPK color scheme? Which colors is used for which atom?
- Physical Molecular Models
- Модель Кори-Полинга-Колтуна: стандарт «молекулярного конструктора»
Источник
МОДЕЛИ ПОЛИПЕПТИДОВ ПОЛИНГА И КОРИ [c.21]
Вскоре появляется знаменитая серия работ Л. Полинга и Р. Кори (1951 г.), в которых авторы рассмотрели все ранее известные структурные модели полипептидов, в том числе предложенные Брэггом, Кендрью и Перутцем, и отвергли их. Вместо них они предложили две новые низкоэнергетические регулярные пространственные формы — а-спираль и р-складчатый лист. Один виток а-спирали включает 3,6 аминокислотных [c.70]
Первое краткое сообщение о результатах исследования Полингом и Кори пространственной структуры полипептидов и белков появилось в ноябре 1950 г. [57]. В апреле следующего года в одном номере журнала было опубликовано сразу восемь работ Полинга и Кори с подробным изложением полученных результатов, а вскоре появились еще четыре их работы [58—65]. Они сразу же обратили на себя внимание научной общественности, вызвали огромный резонанс и оказали сильное влияние на последующее развитие молекулярной биологии и прежде всего исследований пространственной структуры пептидов и белков. В связи с чем вполне обоснованно разделить исследования, проводимые в этой области, на работы до 1951 г. и последующего периода. Читая какой-либо труд, посвященный структуре пептидов, можно, не зная даты публикации, почти наверняка определить, написан ли он до или после появления в печати работ этих ученых. Исследования Полинга и Кори (1951 г.) имеют теоретический характер. Сделанные авторами предсказания возможных структур полипептидной цепи основаны на следующих постулатах 1) приняты одинаковые значения для длин связей и валентных углов всех пептидных групп полипептидной цепи. В литературе они получили название геометрических параметров Полинга— Кори 2) пептидная группа считалась плоской. Возможны две плоские конфигурации группы, отличающиеся взаимным расположением связей N—Н и С=0, цис- и трамс-переход между ними связан с преодолением высокого потенциального барьера (-20 ккал/моль). При построении моделей Полинг и Кори отдали предпочтение транс-конфигурации пептидной группы. По оценке Р. Кори и Дж. Донахью, отклонение от плоского строения группы на 10° вызывает повышение энергии приблизительно на 1,5, а на 30° — на 6 ккал/моль [66] 3) предполагалась полная насыщенность полипептидной цепи водородными связями. Для водородной связи N—Н…О = С были приняты следующие геометрические и энергетические оценки расстояние N…0 считалось равным 2,8 А, максимальное отклонение от линейности N—Н…0 не должно превышать 30° и энергия связи — 8,0 ккал/моль 4) при построении моделей пептидной цепи выбирались наиболее благоприятные ориентации пептидных групп, разделенных атомом С , с учетом потенциалов внутреннего вращения вокруг связей С —N и С —С и ван-дер-ваальсовых контактов между атомами 5) конформационные состояния всех звеньев пептидной цепи считались эквивалентными. [c.21]
Кори нашли, что все они не удовлетворяют приведенным требованиям и должны быть отвергнуты. Авторы предложили две новые структуры (а и у) и структуру складчатого листа (Р). Как известно, до работ Полинга и Кори было разработано большое число молекулярных моделей полипептидов, а Брэгг, Кендрью и Перутц проанализировали этот вопрос в общем виде и как будто предусмотрели все возможные варианты. Как же в таких случаях удалось предложить совершенно новые структуры Это объясняется двумя обстоятельствами. Одно из них связано с более жесткими требованиями, предъявленными Полингом и Кори к геометрии полипептидов другое, сыгравшее решающую роль, — с предположением о реализации спиралей с нецелочисленными винтовыми осями симметрии. Во всех предпринимавшихся ранее поисках структур полипептидов использовались элементы симметрии атомных и низкомолекулярных кристаллов. В этом случае винтовые оси, действительно, могут быть только целочисленными. По отношению к отдельной макромолекуле требование, чтобы спираль содержала только целое число аминокислотных остатков на виток, не может быть оправдано соображениями физического порядка. Использование до Полинга и Кори целочисленных винтовых осей — результат автоматического перенесения кристаллографического принципа симметрии на спиральные полимерные объекты, т.е. в область, где он не работает. [c.22]
В первых моделях пространственного строения белковых молекул водородная связь служила главным стабилизирующим фактором. Молекулы фибриллярных белков представлялись слоистыми структурами, а молекулы глобулярных белков — пакетом слоистых структур, состоящих или из плоских пептидных цепей, скрепленных водородными связями, или из свернутых, регулярных цепей, также стабилизированных внутренними водородными связями. Отклонение от регулярности, повороты цепей и нарушение оптимальных условий образования внутримолекулярных водородных связей допускались лишь в местах аминокислотной последовательности, где находятся остатки пролина. Среди тех, кто считал водородную связь не только главной, но и единственной упаковочной силой, был М. Хаггинс [10, 11]. Все предложенные им структуры полипептидов были полностью насыщены водородными связями. Этот же принцип был положен в основу конструирования пептидных моделей Л. Брэггом, Дж. Кендрью и М. Перутцем [12], а также а-спирали и -структуры складчатых листов Л. Полингом и Р. Кори [1,2]. [c.233]
Может ли полипептидная цепь быть уложена в структуру, состоящую из регулярно повторяющихся участков Чтобы ответить на этот вопрос, Полинг и Кори сравнили ряд потенциально возможных конформаций полипептидов, построив их точные молекулярные модели. При этом строго соблюдались экспериментально установленные для аминокислот и небольших пептидов величины углов связей и межатомных расстоя= ний. В 1951 г. они предложили две периодические полипептидные структуры, названные соответственно о -спираль и р-складчатый слой. [c.34]
Под конформацпоннымн превращениями в макромолекулах до самого недавнего времени понимали превращения (переходы) спираль — клубок в полипептидах и нуклеиновых кислотах. Предполагалось, что, в отличие от макромолекул нативных белков, нуклеиновых кислот и их синтетических моделей — полипептидов и полинуклеотидов, где внутримолекулярные взаимодействия (в основном, водородные связи) обеспечивают наличие вторичной структуры, внутримолекулярные силы у обычных синтетических поли.меров недостаточны для поддержания уиорядоченности в цепи. Макро.молекулы первых существуют в растворах в конформации одионитевых (белки, полипептиды) или двунитевых (нуклеиновые кислоты, полинуклеотиды) спиралей (см. [251, 510]). Двойная спираль Крика — Уотсона [511] для дезоксирибонуклеиновой кислоты и а-сиираль Полинга — Кори [512] для полипептидов — наиболее известные примеры вторичной молекулярной структуры. Макромолекула в спиральной конформации подобна по своей структуре одномерному кристаллу. Изменением температуры или других условий (состав смешанного растворителя, pH растворителя — [c.252]
При поиске решения структурной проблемы белка особенно вдохновляющими примерами явились результаты теоретических исследований Л. Полинга и Р. Кори регулярных структур полипептидов [53] и Дж. Уотсона и Ф. Крика двойной спирали ДНК [54]. В этих работах с помощью простейшего варианта конформационного анализа — проволочных моделей, получивших позднее название моделей Кендрью-Уотсона, а также ряда экспериментальных данных, прежде всего результатов рентгеноструктурного анализа волокон (в случае ДНК еще и специфических соотношений оснований Э. Чаргаффа), удалось предсказать наиболее выгодные пространственные структуры полимеров. Собственно, предсказана была как в случае пептидов, так и нуклеиновых кислот, геометрия лишь одного звена, которое в силу регулярности обоих полимеров явилось трансляционным элементом. Белок же — гетерогенная аминокислотная последовательность, и поэтому таким путем предсказать его трехмерную структуру нельзя. Но то обстоятельство, что простейший, почти качественный, конформационный анализ привел к количественно правильным геометрическим параметрам низкоэнергетических форм звеньев, повторяющихся в гомополипептидах и ДНК, указывало на большие потенциальные возможности классического подхода и его механической модели в описании пространственного строения молекул. [c.108]
На рис. 54 приведены данные по оптическому вращению (для О-линии натрия) олигомеров у-метил-Ь-глутамата в четырех растворителях. Поведение полипептида в дихлоруксусной кислоте — растворителе, способствующем образованию конформации статистического клубка,— такое, которого следует ожидать для конфигурационной оптической активности, т. е. вращение постепенно приближается к асимптоте, когда п очень велико. Было обнаружено, что для нескольких изученных высокомолекулярных поли-у-метил-Ь-глутаматов средняя величина [а] приблизительно равна —33°, что очень близко к ожидаемой предельной величине [0)1) на рис. 54, а (Доти и Янг, неопубликованные данные). Совершенно другая картина на рисунках 54, б—г, на которых удельное вращение проходит через минимум, соответствующий пентамеру, и затем становится положительным при высоких СП. Из ранних работ Доти, Блоута с сотр. [30, 31 ] известно, что диметилформамид, л-крезол и диоксан — растворители, способствующие образованию конформации а-спирали. В соответствии с моделью а-спирали, предложенной Полингом и Кори, на виток спирали приходится 3,6 остатка и первая водородная связь образуется внутримолекулярно между первым и пятым остатками. Таким образом, результаты, приведенные на рис. 54, можно интерпретировать в терминах образования а-спирали для пентамера и высших гомологов. О диоксане также известно, что он вызывает ассоциацию молекул и появление так называемой Р-формы у полипептидов низкого молекулярного веса (раздел Г-7). Гудман и др. пришли к выводу о том, что данные, приведенные на рисунке 54, г, являются результатом одновременного влияния внутримолекулярных водородных связей и межмолекулярной ассоциации. [c.101]
В 1950 г., за несколько месяцев до опубликования серии исследований Л. Полинга и Р. Кори, появилась большая работа Л. Брэгга, Дж. Кендрью и М. Перутца [56], которая как бы подвела черту под структурными исследованиями белков и полипептидов начального периода. В этой работе анализируются структурные модели Астбери, Хаггинса, Цана и других авторов и предлагается ряд новых моделей. Однако, по-прежнему, речь идет о возможных типах укладки основной пептидной цепи. Авторы также рассматривают свои первые результаты рентгеноструктурного изучения гемоглобина и миоглобина. При анализе возможных структур пептидного остова Брэгг, Кендрью и Перутц принимают геометрические параметры пептидной группы, близкие к параметрам Хаггинса и согласующиеся с более поздними данными Кори. Так, длина связи С -С считается равной 1,52 А, С -М — 1,36 А и С =0 — 1,24 А. Валентные углы при атоме С приняты тетраэдрическими (109°28 ), а при атоме N — или тетраэдрическими, или тригональными (120°). Длина водородной связи М-Н…С=0 составляет 2,85 А. Ее направленность строго не лимитируется, но авторы по мере возможности стремятся к линейному расположению связей К-Н и С=0. Л. Брэгг, Дж. Кендрью и М. Перутц приняли постулат М. Хаггинса о спиральной симметрии полипептидной цепи с целочисленным порядком винтовой оси, но отвергли его второй постулат о том, что каждый элемент основной цепи должен находиться в одинаковых отношениях с соседними элементами (принцип эквивалентности). Так же как и Хаггинс, они придают огромное значение водородной связи М-Н…С = 0 в формировании структуры основной цепи и считают наиболее стабильными только те из них, которые полностью насыщены ими. Предложенная авторами классификация спиральных структур является общепризнанной и в настоящее время [56]. В основу ее положено разделение структур по симметрии пептидного остова и размеру циклов с водородными связями. [c.19]
Картер и Краут показали, что весьма распространенная вытянутая конформация полипептидных цепей (р-складчатая структура по Полингу и Кори) соответствует стабильной правозакрученной двойной спирали, т. е. существует двойная полипептидная спираль. Ее шаг и радиус примерно такие же, как и у двойной спирали нуклеиновых кислот. Авторы весьма тщательно построили пространственные молекулярные модели и увидели, что полипептидная двойная спираль в точности комплементарна двойной спирали РНК, причем полипептидная двойная спираль точно входит в малую бороздку двойной спирали РНК. В результате образуются водородные связи между 2 -0Н Группами рибозы (что, по-видимому, и объясняет особые свойства РНК, у ДНК нет этого гидроксила) и кислородом карбонила пептидной связи. Авторы отмечают, что такую точную комплементарность ста- бильных конформаций РНК и полипептидов вряд ли можно считать простым совпадением. Они предположили, что обе эти двойные спирали взаимно катализировали сборку друг друга из активированных предшественников на ранних стадиях эволюции. В самом деле, комплементарное соответствие двух двойных спиралей представляется весьма важным, но оно обеспечивает ускорение синтеза лишь основных каркасов — полипептидного и полинуклеотидного, тогда как для преодоления обсуждаемого нами кризиса на начальной стадии биологической эволюции необходимо установление полного однозначного соответствия определенных аминокислотных и нуклеотидных радикалов. Для этого нужно, чтобы взаимная полимеразная активность поли- [c.59]
Источник

ПЕРВИЧНАЯ СТРУКТУРА БЕЛКА
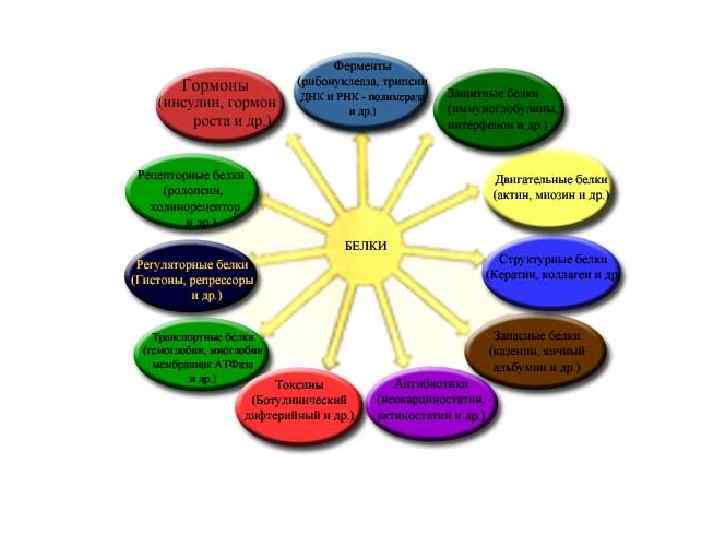

БЕЛКИ – БИОПОЛИМЕРЫ, МОНОМЕРАМИ КОТОРЫХ ЯВЛЯЮТСЯ АМИНОКИСЛОТЫ. РАЗЛИЧАЮТ L и D формы аминокислот БЕЛКИ СОСТОЯТ ТОЛЬКО ИЗ L АМИНОКИСЛОТ

АМИНОКИСЛОТЫ, ВХОДЯЩИЕ В СОСТАВ БЕЛКА
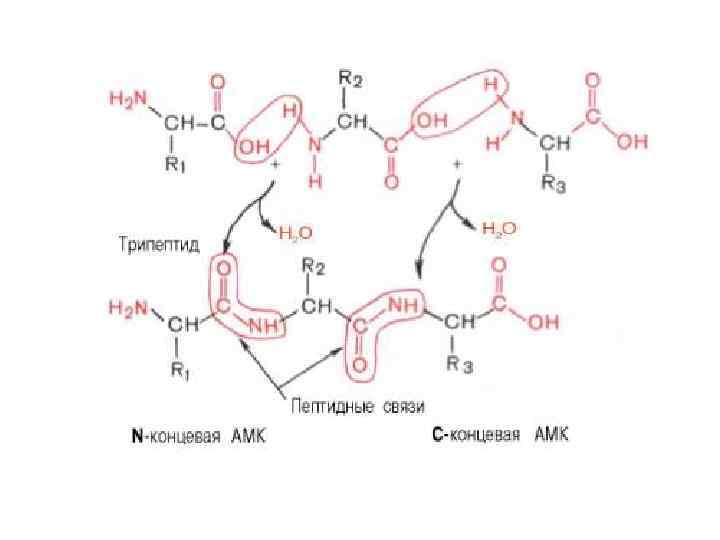

ПЕРВИЧНАЯ СТРУКТУРА БЕЛКА

ДЕЛОКАЛИЗАЦИЯ -ЭЛЕКТРОНОВ В ПЕПТИДНОЙ СВЯЗИ ЭЛЕКТРОНЫ АТОМОВ СИ N СОЗДАЮТ «ДЕЛОКАЛИЗОВАННОЕ » ЭЛЕКТРОННОЕ ОБЛАКО, КОТОРОЕ ОХВАТЫВАЕТ ОБА ЭТИ АТОМА И РАСПРОСТРАНЯЮЩЕЕСЯ НА АТОМ О ЧЕРЕЗ СВЯЗЬ С=О. ОБОБЩЕСТВЛЕННЫЕ ЭЛЕКТРОНЫ ПРЕПЯТСТВУЮТ ВРАЩЕНИЮ ВОКРУГ C-N СВЯЗИ

ПРИ ДЕЛОКАЛИЗАЦИИ ЭНЕРГИЯ ЭЛЕКТРОНОВ ПАДАЕТ И ОНИ ПЕРЕХОДЯТ В БОЛЕЕ СТАБИЛЬНОЕ СОСТОЯНИЕ
 0, 132 НМ N – C (ОДИНОЧНАЯ)")
ДЛИНЫ СВЯЗЕЙ N – C (ПЕПТИДНАЯ СВЯЗЬ) 0, 132 НМ N – C (ОДИНОЧНАЯ) 0, 147 НМ N = C (ДВОЙНАЯ) 0, 125 НМ 0, 147

ДЛИНЫ СВЯЗЕЙ В ПЕПТИДНОЙ ГРУППЕ H H 0, 132 нм 0, 147 нм С : N С 0, 124 нм 0, 121 нм O O- N+

ПРИ КОМНАТНОЙ ТЕМПЕРАТУРЕ ВАЛЕНТНЫЕ СВЯЗИ НЕ ФЛУКТУИРУЮТ: ЧАСТОТА КОЛЕБАНИЙ ЭТИХ СВЯЗЕЙ 2 х1013 сек ЧАСТОТА ТЕПЛОВЫХ КОЛЕБАНИЙ 7 х1012 сек – 1 АМПЛИТУДА ТЕПЛОВЫХ КОЛЕБАНИЙ ВАЛЕНТНЫХ УГЛОВ НЕВЕЛИКА И СОСТАВЛЯЕТ ОКОЛО 5 о. ТАКИМ ОБРАЗОМ, ГИБКОСТЬ ПОЛИПЕПТИДНОЙ ЦЕПИ ОБЕСПЕЧИВАЕТСЯ ДРУГИМИ ФАКТОРАМИ.

ГИБКОСТЬ ПОЛИПЕПТИДНОЙ ЦЕПИ, А, ЗНАЧИТ, И ЕЕ СПОСОБНОСТЬ ОБРАЗОВЫВАТЬ СПИРАЛИ И ГЛОБУЛЫ ОБЕСПЕЧИВАЕТ ВРАЩЕНИЕ ВОКРУГ ВАЛЕНТНЫХ СВЯЗЕЙ.

УГОЛ ОПИСЫВАЕТ ВРАЩЕНИЕ ВОКРУГ СВЯЗИ N – C УГОЛ ОПИСЫВАЕТ ВРАЩЕНИЕ ВОКРУГ СВЯЗИ C – C УГОЛ ОПИСЫВАЕТ ВРАЩЕНИЕ ВОКРУГ СВЯЗИ C — N — УГОЛ ВРАЩЕНИЯ В БОКОВОЙ ЦЕПИ
 МАЛОВЕРОЯТЕН ИЗ-ЗА ЕЕ ЧАСТИЧНО ДВОЙНОГО ХАРАКТЕРА. ЗАТО ВОЗМОЖНЫ")
ПОВОРОТ ВОКРУГ СВЯЗИ С-N (УГОЛ ) МАЛОВЕРОЯТЕН ИЗ-ЗА ЕЕ ЧАСТИЧНО ДВОЙНОГО ХАРАКТЕРА. ЗАТО ВОЗМОЖНЫ ПОВОРОТЫ ВОКРУГ ОДИНАРНЫХ СВЯЗЕЙ C -C (УГОЛ ) И C -N (УГОЛ ) ЕА ДЛЯ УГЛОВ И ОТ 0, 84 ДО 2, 52 к. Дж/моль, ДЛЯ ПЕПТИДНОЙ СВЯЗИ 84 к. Дж/моль
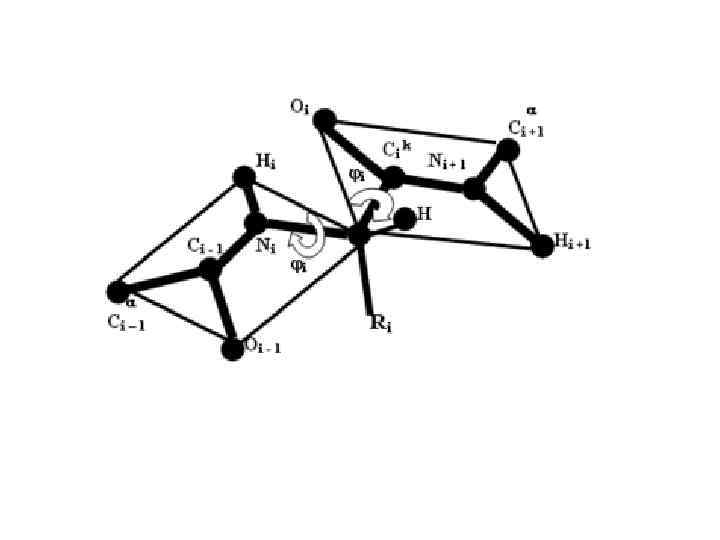

ВТОРИЧНАЯ СТРУКТУРА БЕЛКА
ГЛОБУЛА

НАИБОЛЕЕ ВАЖНЫМИ ДЛЯ ВТОРИЧНОЙ СТРУКТУРЫ БЕЛКА ЯВЛЯЮТСЯ ВОДОРОДНЫЕ СВЯЗИ -СКЛАДЧАТАЯ СТРУКТУРА -СПИРАЛЬ Водородные связи параллельны оси спирали Водородные связи перпендикулярны оси спирали

Водородные связи в спирали • параллельны оси спирали • образуются между первым и четвертым аминокислотными остатками
 СПИРАЛИ ПРИХОДИТСЯ 3, 6 АМИНОКИСЛОТНЫХ ОСТАТКОВ. ШАГ СПИРАЛИ")
ПАРАМЕТРЫ -СПИРАЛИ НА КАЖДЫЙ ВИТОК (ШАГ) СПИРАЛИ ПРИХОДИТСЯ 3, 6 АМИНОКИСЛОТНЫХ ОСТАТКОВ. ШАГ СПИРАЛИ (РАССТОЯНИЕ ВДОЛЬ ОСИ) – 0, 54 НМ. НА ОДИН АМИНОКИСЛОТНЫЙ ОСТАТОК ПРИХОДИТСЯ 0, 15 НМ. ЧЕРЕЗ 5 ВИТКОВ СПИРАЛИ (18 АМИНОКИСЛОТНЫХ ОСТАТКОВ) СТРУКТУРНАЯ КОНФИГУРАЦИЯ ПОЛИПЕПТИДНОЙ ЦЕПИ ПОВТОРЯЕТСЯ

ПОСТУЛАТЫ ПОЛИНГА – КОРИ • Длины связей и валентных углов всех пептидных групп одинаковы • Полипептидная цепь насыщена водородными связями • Конформационное состояние всех звеньев полипептидной цепи является эквивалентным

-складчатые структуры

АНТИПАРАЛЛЕЛЬНЫЕ И ПАРАЛЛЕЛЬНЫЕ -СКЛАДЧАТЫЕ СТРУКТУРЫ

-СКЛАДЧАТАЯ СТРУКТУРА Водородные связи перпендикулярны оси спирали

В БЕЛКАХ, КРОМЕ ТОГО, МОЖНО ОБНАРУЖИТЬ НЕУПОРЯДОЧЕННЫЕ УЧАСТКИ: ГЕМОГЛОБИН: ПОЛИПЕПТИДНЫЕ ЦЕПИ СПИРАЛИЗОВАНЫ НА 75% ТРИПСИН: ПОЛИПЕПТИДНАЯ ЦЕПЬ СПИРАЛИЗОВАНА НА 30%
 СПИРАЛИ •")
ДРУГИЕ СПИРАЛЬНЫЕ КОНФОРМАЦИИ СПИРАЛЬ 310 • 3 ОСТАТКА АМИНОКИСЛОТ НА ШАГ (ВИТОК) СПИРАЛИ • РАДИУС 0, 19 НМ p. СПИРАЛЬ 4, 4 ОСТАТКА НА ШАГ СПИРАЛИ РАДИУС 0, 28 НМ

СВЕРХВТОРИЧНЫЕ СТРУКТУРЫ

СВЕРХВТОРИЧНЫЕ СТРУКТУРЫ – ЭТО ТЕРМОДИНАМИЧЕСКИ ИЛИ КИНЕТИЧЕСКИ СТАБИЛЬНЫЕ КОМПЛЕКСЫ -СПИРАЛЕЙ И СТРУКТУР, КОТОРЫЕ ФОРМИРУЮТСЯ ЗА СЧЕТ МЕЖРАДИКАЛЬНЫХ ВЗАИМОДЕЙСТВИЙ.

Примеры суперспиралей Легкий меромиозин и тропомиозин – 2 скрученные спирали

СУПЕРСПИРАЛЬ, СОСТОЯЩАЯ ИЗ НЕСКОЛЬКИХ -СПИРАЛЕЙ В ПЕРЕВИТЫХ СПИРАЛЯХ ПЕРИОДИЧНОСТЬ 3, 5 ОСТАТКОВ НА ВИТОК (ВМЕСТО 3, 6 В ОДИНОЧНОЙ -СПИРАЛИ )
 и тройной (б) суперспирали (вид с торца спирали). В")
Взаимодействие a-спиралей в двойной (а) и тройной (б) суперспирали (вид с торца спирали). В двойной суперспирали непосредственно контактируют с другой спиралью только остатки а и d, а в тройной — еще и остатки e и g (хотя и более слабо).

МЕТОДЫ ИЗУЧЕНИЯ ВТОРИЧНОЙ СТРУКТУРЫ БЕЛКА

ДИСПЕРСИЯ ОПТИЧЕСКОГО ВРАЩЕНИЯ и КРУГОВОЙ ДИХРОИЗМ – методы, основанные на взаимодействии поляризованного света с оптически активными молекулами

Метод КРУГОВОГО ДИХРОИЗМА основан на различии в поглощении право- и левополяризованного света оптически активным веществом.

Левый и правый поляризованные по кругу лучи поразному поглощаются средой, т. е где коэффициенты экстинкции для лучей с левой и правой круговой поляризацией. Суммирование соответствующих им ВЕКТОРОВ неравной величины El и Еr дает результирующий вектор, конец которого описывает эллипс, т. е. плоскополяризованный свет после прохождения через оптически активную среду становится эллиптически поляризованным. Это явление называют КРУГОВЫМ ДИХРОИЗМОМ.

ПАРАМЕТРЫ, ОПРЕДЕЛЯЕМЫЕ МЕТОДОМ КРУГОВОГО ДИХРОИЗМА
![Молярная эллиптичность [ ] Характерные формы спектров КД для полилизина в форме aспирали (a),](https://present5.com/presentation/53476435_171788473/image-37.jpg "Молярная эллиптичность [ ] Характерные формы спектров КД для полилизина в форме aспирали (a),")
Молярная эллиптичность [ ] Характерные формы спектров КД для полилизина в форме aспирали (a), b-структуры (b) и неупорядоченного клубка (r).

ДИСПЕРСИЯ ОПТИЧЕСКОГО ВРАЩЕНИЯ Оптически активное вещество характеризуется разными показателями преломления n. L и n. R для лево- и правовращающих компонент света, в результате чего происходит вращение плоскости поляризации.

ПАРАМЕТРЫ, ОПРЕДЕЛЯЕМЫЕ МЕТОДОМ ДОВ Оптическое вращение Удельное оптическое вращение Молярное вращение
. Спектры")
В белках спектры ДОВ и КД регистрируют в области поглощения пептидных групп (УФ-область). Спектры КД будут отличаться в спиралях различной закрученности.

МЕТОДЫ ИЗУЧЕНИЯ ДИНАМИКИ БЕЛКОВЫХ СТРУКТУР – ЯМР ЭПР И

ЭЛЕКТРОННЫЙ ПАРАМАГНИТНЫЙ РЕЗОНАНС

Метод электронного парамагнитного резонанса был открыт в 1944 г. в Казанском Университете Е. К. ЗАВОЙСКИМ. Пионерами применения ЭПР в биологических исследованиях в России были Л. А. Блюменфельд и А. Э. Калмансон (1958).

УСЛОВИЕ ВОЗНИКНОВЕНИЯ ЭПР – НАЛИЧИЕ У ЧАСТИЦ НЕСКОМПЕНСИРОВАННЫХ МАГНИТНЫХ МОМЕНТОВ, ОБУСЛОВЛЕННЫХ СПИНОМ НЕСПАРЕННЫХ ЭЛЕКТРОНОВ
.")
ЭФФЕКТ ЗЕЕМАНА В отсутствии внешнего магнитного поля магнитные моменты электронов ориентированы случайным образом (А). При наложении внешнего магнитного поля магнитные моменты электронов ориентируются в поле в зависимости от величины спинового магнитного момента (Б).

УСЛОВИЕ РЕЗОНАНСА — разница энергии между уровнями H — напряженность магнитного поля — магнетон Бора g – фактор спектроскопического расщепления

На систему электронов, находящуюся в магнитном поле, подают электромагнитную энергию, при определенных значениях величины энергии падающего кванта будут происходить переходы электронов между уровнями. Необходимым условием переходов является равенство энергии падающего кванта (h ) и разности энергий между уровнями электронов с различными спинами (gb. H).
 = const d. I/d. H Первая производная d. I/d. H")
СПЕКТРЫ ЭПР Iпогл =f(H) = const d. I/d. H Первая производная d. I/d. H

ИСПОЛЬЗОВАНИЕ ЭПР ДЛЯ БЕЛКОВ – МЕТОД СПИНОВЫХ МЕТОК. В ИХ РОЛИ – НИТРОКСИЛЬНЫЕ РАДИКАЛЫ Формула спинового зонда ТЕМПО и его спектр ЭПР. В этом радикале неспаренный электрон локализован около ядра азота, магнитное поле которого накладывается на внешнее магнитное поле.

ЯДЕРНЫЙ МАГНИТНЫЙ РЕЗОНАНС
 – резонансное поглощение электромагнитной энергии веществом, обусловленное переориентацией магнитных моментов")
Ядерный магнитный резонанс (ЯМР) – резонансное поглощение электромагнитной энергии веществом, обусловленное пере
